北京时间2021年10月27日,国际学术期刊Nature自然杂志在线发表了十大网投官方入口-加州大学河滨分校联合中心(FAFU-UCR联合中心)关于植物“酸性生长理论”的研究文章。揭示了植物激素生长素通过细胞膜表面类受体激酶家族蛋白(TMK),激活膜上质子泵,酸化细胞壁,促进细胞的伸长及组织生长的重要分子机制。我司林文伟教授为论文第一作者,加州大学河滨分校杨贞标教授为论文通讯作者。十大网投官方入口徐通达教授、日本名古屋大学Toshinori Kinoshita和明尼苏达大学William Gray参与了该研究工作。

细胞伸长是动植物生长的基础。对于包埋于坚固细胞壁中的植物细胞来说,要扩展和伸长,首先需要通过修饰和调节细胞壁的理化性质,松弛细胞壁,提高其延展性,在此基础上细胞胞内膨压才能驱动细胞伸展。近百年的研究发现,植物激素生长素 (auxin)是调控细胞壁动态变化的关键元件。生长素(auxin)是最早被发现的植物激素,因其促进生长的核心功能取名于希腊语“auxein”即”生长“的意思。 生长素作为植物中最重要的激素之一,几乎参与了植物所有的生长发育过程以及对复杂环境适应过程。而其行使功能的关键在于对细胞生长的精准调控。早在1971年,著名科学家 Achim Hager 和 Robert Cleland 首次提出了细胞的“酸性生长假说” (acid-growth hypothesis): 生长素通过细胞膜质子泵ATP酶(PM H+-ATPase, AHA)的激活引起质外体(Apoplast, 介于细胞膜与细胞壁之间)酸化,激活相关的蛋白酶修饰细胞壁, 提高细胞壁的延展性,在细胞内膨压驱使下,促进细胞伸长(图1)。然而半个世纪以来,该假说的具体机制仍然未知。
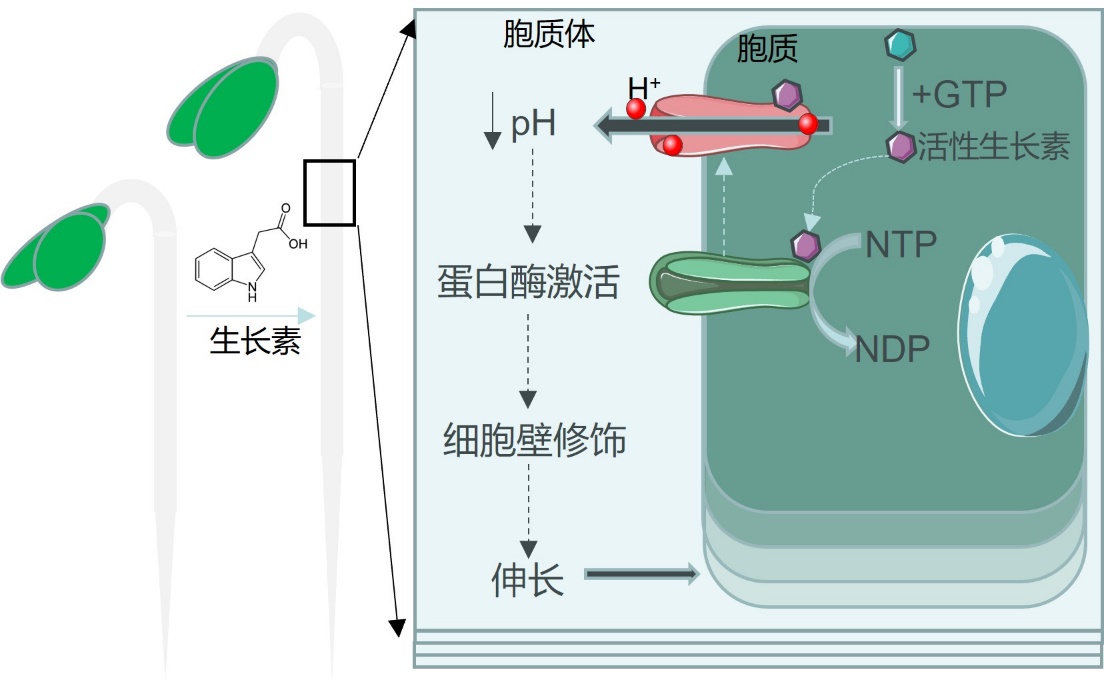
图1. Hager 等(1971)第一次提出的酸性生长假说示意图
随着分子生物学的不断发展,利用新的研究手段和方法,科学家们不断地挑战或者佐证植物细胞的“酸性生长假说”,却一直未有定论,其核心问题在于“生长素如何激活定位在细胞膜上的质子泵蛋白H+-ATPase的活性,引起质子外流”。 已有研究结果表明生长素通过经典的TIR1/AFB-Aux/IAA-ARF信号通路,间接地诱导H+-ATPase碳末端保守的苏氨酸位点(AHA2, T947)磷酸化激活质子泵活性,降低细胞壁的pH值,介导了拟南芥下胚轴的伸长。然而生长素是通过何种激酶直接磷酸化修饰来激活质子泵蛋白的活性,成为了该假说最迫切需要回答的问题。
加州大学河边分校杨贞标团队以及十大网投官方入口徐通达团队前期的研究表明,类受体激酶家族TMK蛋白(Transmembrane Kinase)介导的生长素信号通路在植物各个生长发育过程中发挥着至关重要的作用,从非转录水平到转录水平多维度调控植物细胞对生长素的响应。基于这些前期基础,通过免疫沉淀结合蛋白质谱的方法鉴定到TMK1特异性地结合定位于细胞膜上的质子泵家族成员(H+-ATPase, AHAs)。利用微流控技术结合实时荧光能量共振转移实验证明,生长素可以在数秒内,诱导TMK1和 AHA1在细胞膜上结合,形成蛋白复合体(图2A-B)。 进一步研究发现,生长素直接通过TMK激酶特异性磷酸化质子泵蛋白AHA碳末端保守的苏氨酸位点(AHA2, T947), 激活其质子泵活性,导致大量的质子被泵出细胞外,从而引起细胞壁酸性化和细胞的伸长(图2C-D)。
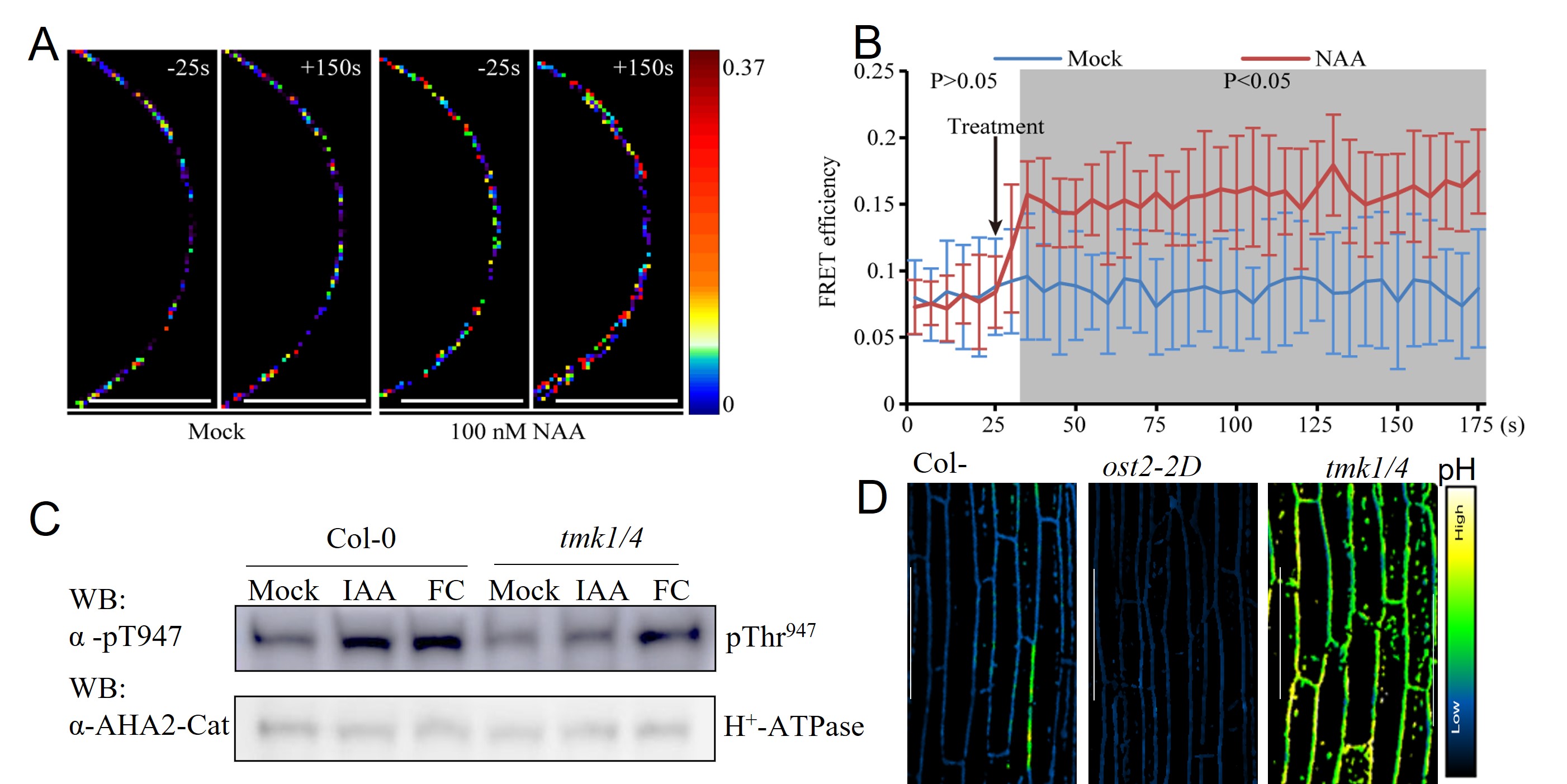
图2. 生长素通过TMK磷酸化并激活质子泵(H+-ATPase)导致细胞壁酸化和伸长
拟南芥下胚轴生长是研究细胞伸长的经典模型,研究人员发现在TMK 缺失突变体有显著的下胚轴伸长缺陷,且生长素无法激活质子泵蛋白活性,导致突变体中整体的质子泵活性显著低于野生型水平, 细胞壁pH升高导致细胞壁无法软化,从而抑制了细胞和组织的伸长(图2D)。通过激活质子泵蛋白活性或者置于酸性生长环境,都可以在一定程度上的促进TMK缺失突变体的下胚轴伸长,进一步证明了TMK介导的细胞壁酸性化是生长素促进细胞生长的关键机制。
最终,该研究解析了基于TMK介导的生长素信号通路促进细胞酸性生长的新机制,为半世纪以来一直未解的 “酸性生长假说“ 提供直接理论依据,是植物细胞生长调控的突破性进展,在植物领域也具有广泛的应用前景(图3)。
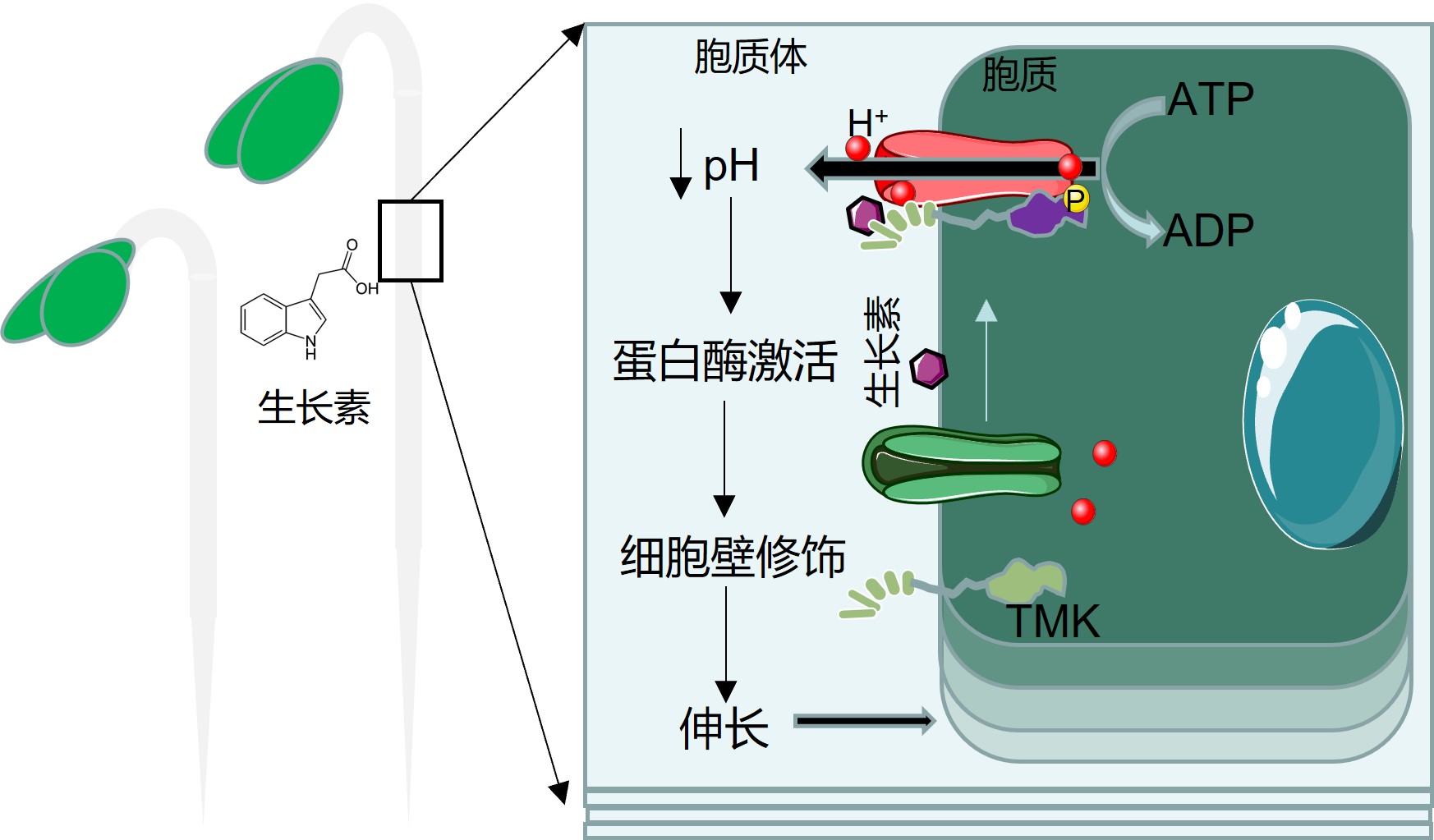
图3. TMK调控生长素介导的细胞“酸性生长”

